Εισαγωγή
Με την αύξηση του παγκόσμιου πληθυσμού και τη συνεχώς αυξανόμενη ζήτηση πρωτεϊνών, ορισμένες εναλλακτικές πηγές πρωτεϊνών έχουν προσελκύσει πρόσφατα αυξανόμενο ενδιαφέρον, όπως οι πρωτεΐνες φυτικής προέλευσης, οι πρωτεΐνες κρέατος που καλλιεργούνται ή καλλιεργούνται, οι πρωτεΐνες που παράγονται από ζύμωση, οι πρωτεΐνες βρώσιμων εντόμων και τα φύκια [1]. Η πηκτική ιδιότητα είναι από τις σημαντικότερες λειτουργίες των εναλλακτικών πρωτεϊνών, συμβάλλοντας στην υφή και τη γεύση των τροφίμων. Η ζελατινοποίηση συμβαίνει κατά τη διάρκεια της επεξεργασίας και της παρασκευής των τροφίμων. Η θέρμανση είναι μία από τις πιο συχνά χρησιμοποιούμενες μεθόδους για τον σχηματισμό πηκτωμάτων με εναλλακτικές πρωτεΐνες. Μετά τη μετουσίωση και το ξεδίπλωμα των πρωτεϊνικών μορίων με θέρμανση και την περιβολή νερού, αυτά θα συσσωματωθούν για να σχηματίσουν μια τρισδιάστατη δομή δικτύου, δηλαδή τη δομή πηκτής.
Στην πράξη, ένα περιστροφικό ρεόμετρο είναι κατάλληλο για τη μελέτη των ιδιοτήτων πηκτωματοποίησης των εναλλακτικών πρωτεϊνών που προκαλούνται από τη θερμότητα, όπως η θερμοκρασία πηκτωματοποίησης, η σταθερότητα της πηκτής και η αντοχή της πηκτής.
Υλικά και συνθήκες μέτρησης
Οι πρωτεϊνικές σκόνες διασκορπίστηκαν σε απομεταλλωμένο νερό σε καθορισμένες συγκεντρώσεις πρωτεϊνών (συμπύκνωμα πρωτεϊνών μπιζελιού: 10 wt% και 7 wt%, πρωτεΐνες εντόμων: 10 wt%). Τα εναιωρήματα πρωτεϊνών αναδεύτηκαν με μαγνητικό αναδευτήρα σε θερμοκρασία δωματίου για 2 ώρες. Εκτός από το δείγμα ολόκληρου του αυγού, παρασκευάστηκε και δείγμα ασπράδι αυγού με αφαίρεση του κρόκου και έντονο χτύπημα με σύρμα σε θερμοκρασία δωματίου για λίγα λεπτά ώστε να προκύψει ομοιογενές διάλυμα.
Για τη διεξαγωγή των μετρήσεων στα δείγματα συμπυκνωμένης πρωτεΐνης μπιζελιού, στο ολόκληρο αυγό και στα δείγματα ασπράδι αυγού χρησιμοποιήθηκε ρεόμετρο NETZSCH Kinexus Prime pro+ , εξοπλισμένο με σύστημα πλάκας-πλάκας (διάμετρος: 40 mm, διάκενο: 0,5 mm). Πραγματοποιήθηκε σάρωση θερμοκρασίας με αύξηση της θερμοκρασίας από 25°C έως 95°C με ρυθμό 5°C/min. Μετά την επίτευξη της υψηλότερης θερμοκρασίας, οι πηκτές που προκλήθηκαν από τη θερμότητα παρέμειναν για 10 λεπτά για να μελετηθεί η σταθερότητα της πηκτής. Το μέτρο αποθήκευσης (G') και το μέτρο απώλειας (G'') καταγράφηκαν κατά τη διάρκεια του πειράματος. Τα αποτελέσματα που προέκυψαν για τις πρωτεΐνες φυτικής προέλευσης και τις πρωτεΐνες αυγών συγκρίθηκαν με εκείνα για τις πρωτεΐνες εντόμων [2].

Μέση: Φρέσκα αυγά (που χρησιμοποιήθηκαν ως ζωικές πρωτεΐνες για σύγκριση): αναλύθηκαν τόσο το ολόκληρο αυγό όσο και το ασπράδι αυγού, αντίστοιχα.
Δεξιά: Νέες πρωτεΐνες μη φυτικής προέλευσης: πρωτεΐνες εντόμων (περιεκτικότητα σε ακατέργαστη πρωτεΐνη 68,7 g/100 g) που εξήχθησαν από έντομα κουκουλιών μύγας μαύρου στρατιώτη (BSF) σε προηγούμενη μελέτη [2]
Αποτελέσματα μετρήσεων και συζήτηση
Οι πηκτικές ιδιότητες διαφόρων εναλλακτικών πρωτεϊνών και μιας τυπικής πηγής ζωικής πρωτεΐνης (αυγό) μελετήθηκαν κατά τη διάρκεια και μετά τη θέρμανση με περιστροφική ρεομετρία.
Στο Σχήμα 2 παρουσιάζεται το μέτρο αποθήκευσης, G', και το μέτρο απώλειας, G'', της γέλης που σχηματίζεται με συμπύκνωμα πρωτεΐνης μπιζελιού κατά τη διάρκεια θερμικής επεξεργασίας. Παρατηρείται αύξηση των G' και G'' όταν η θερμοκρασία αυξάνεται μέχρι τους 55°C περίπου. Αυτό προκύπτει από τη μετουσίωση της πρωτεΐνης. Μετά τη θερμική επεξεργασία με περαιτέρω αύξηση της θερμοκρασίας, το G' είναι υψηλότερο από το G'' σε συγκέντρωση πρωτεΐνης 10 % κ.β., δείχνοντας συμπεριφορά πηκτής που μοιάζει με στερεό.
Επιπλέον, ένα πείραμα στο συμπύκνωμα πρωτεΐνης μπιζελιού σε χαμηλότερη συγκέντρωση πρωτεΐνης 7 κ.β., δείχνει ότι το G'' είναι υψηλότερο από το G' με την αύξηση της θερμοκρασίας θέρμανσης, υποδηλώνοντας μια ασθενή συμπεριφορά πηκτής που μοιάζει με υγρό.
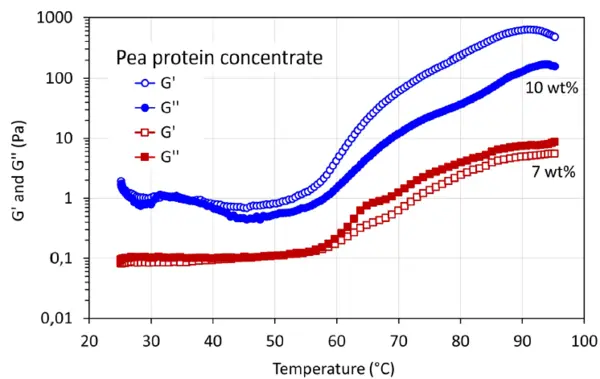
Ωστόσο, η διασταύρωση G' = G'' δεν παρατηρήθηκε στην παρούσα μελέτη συμπυκνωμένης πρωτεΐνης μπιζελιού.
Σε μια προηγούμενη μελέτη της επαγόμενης από τη θερμότητα ζελατινοποίησης με πρωτεΐνες εντόμων κουκουλιών BSF [2], διαπιστώθηκε ότι με την αύξηση της θερμοκρασίας πάνω από τους 50°C, τόσο η G' όσο και η G'' αυξάνονταν, γεγονός που προκλήθηκε από τη μετουσίωση των πρωτεϊνών. Το δείγμα που μελετήθηκε άρχισε να σχηματίζει πηκτή, γεγονός που υποδεικνύεται από τη διασταύρωση G' = G'' στους 60°C, τη θερμοκρασία του σημείου πήξης.
Η εξέλιξη της καμπύλης κατά τη θερμοκρασία για τις πρωτεΐνες των εντόμων είναι διαφορετική από εκείνη του συμπυκνώματος πρωτεϊνών μπιζελιού. Αυτή η διαφορετική συμπεριφορά πηκτωματοποίησης μπορεί να αποδοθεί σε διαφορετικές συνθέσεις υλικών και σε μεμονωμένα χαρακτηριστικά πρωτεϊνών, όπως πιθανά διαφορετικά υδρόφιλα και υδρόφοβα αμινοξέα και οι αναλογίες τους μεταξύ των διαφόρων εναλλακτικών πρωτεϊνών.
Οι καμπύλες ζελατινοποίησης τόσο των δειγμάτων ολόκληρου του αυγού όσο και του ασπράδι αυγού παρουσιάζουν την τυπική μετάβαση sol-gel κατά τη διάρκεια της σάρωσης θερμοκρασίας. Από τους 60°C περίπου παρατηρείται σημαντική αύξηση των G' και G", η οποία εξηγείται, για παράδειγμα, από δομικές αλλαγές ή μετουσίωση των πρωτεϊνών. Το Σχήμα 3 απεικονίζει τα G' και G'' της γέλης που σχηματίζεται με το διάλυμα ολόκληρου του αυγού κατά τη διάρκεια της θερμικής επεξεργασίας. Το G' εμφανίζει εμφανή αύξηση στους 62°C περίπου και απότομη αύξηση στους 75°C περίπου, ενώ το G'' παρουσιάζει δραματική αύξηση στους 75°C περίπου. Το σημείο διασταύρωσης εμφανίζεται στους 74°C περίπου. Για το δείγμα ασπράδι αυγού (Σχήμα 4), τόσο το G' όσο και το G'' εμφανίζουν δύο εμφανείς αυξήσεις στους 64°C περίπου και στους 75°C αντίστοιχα. Το σημείο διασταύρωσης εμφανίζεται περίπου στους 62,5°C. Τα παρατηρούμενα φαινόμενα μετουσίωσης μπορούν να συσχετιστούν με τη χημική σύνθεση του δείγματος ολόκληρου του αυγού (ασπράδι και κρόκος) και του δείγματος ασπράδι.
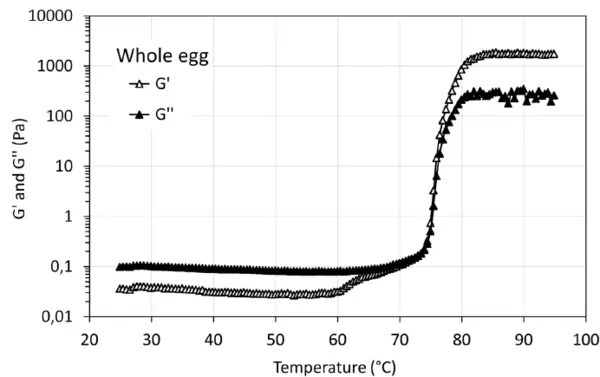
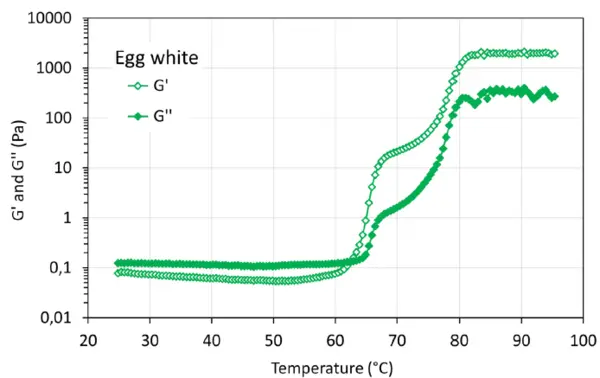
Το Σχήμα 5 απεικονίζει την αντοχή του πηκτώματος, G', και τη σταθερότητα εντός του χρόνου αναμονής 10 λεπτών μετά την επίτευξη της υψηλότερης θερμοκρασίας. Οι πηκτές που προκαλούνται από τη θερμότητα από πρωτεΐνες αυγών παρουσιάζουν την υψηλότερη αντοχή και είναι πολύ σταθερές. Αυτή η σταθερή ιδιότητα πηκτής έχει επίσης παρατηρηθεί σε θερμοκρασία υψηλότερη από 85°C στο σχήμα 3 και στο σχήμα 4. Για το δείγμα συμπυκνωμένης πρωτεΐνης μπιζελιού σε ποσοστό 10 % κ.β., χρειάζονται περίπου 4 λεπτά για να επιτευχθεί η υψηλότερη αντοχή πηκτής και στη συνέχεια η σχηματιζόμενη πηκτή είναι σταθερή, ενώ η αντοχή πηκτής μειώνεται ελαφρώς για το δείγμα συμπυκνωμένης πρωτεΐνης μπιζελιού σε ποσοστό 7 % κ.β.. Αυτό μπορεί να οφείλεται στην παραμόρφωση (καταστροφή) της σχηματιζόμενης αδύναμης δομής πηκτής κατά τη διάρκεια της μέτρησης. Σε σύγκριση με την αντοχή της πηκτής που σχηματίζεται με πρωτεΐνες εντόμων [2], οι εν λόγω πηκτές παρουσιάζουν διαφορετικές αντοχές πηκτής, G', ακολουθώντας τη σειρά
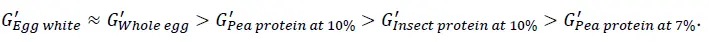
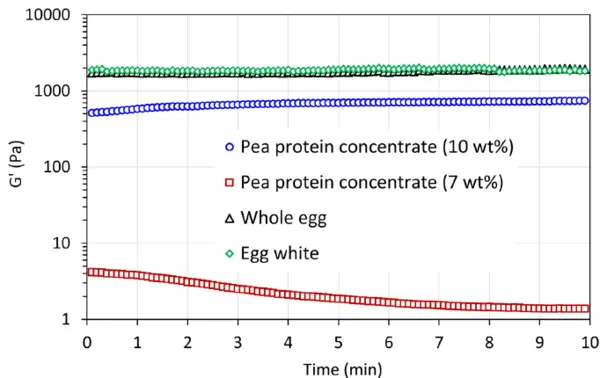
Αυτό μπορεί να υποδηλώνει ότι διαφορετικές εναλλακτικές πρωτεΐνες έχουν διαφορετικές πιθανές εφαρμογές. Για παράδειγμα, οι πηκτές που προκαλούνται από τη θερμότητα με χαμηλότερη τιμή G' ή με ασθενές δίκτυο πηκτής θα μπορούσαν να είναι ενδιαφέρουσες και κατάλληλες για υγρές συνθέσεις τροφίμων, όπως ποτά φυτικής προέλευσης ή εναλλακτικά γάλατα, ενώ οι πηκτές με υψηλότερη τιμή G' ή με ισχυρό δίκτυο πηκτής θα ήταν ενδιαφέρουσες για γαλακτοκομικά και ανάλογα κρέατος κ.ο.κ.
Αξίζει να αναφερθεί ότι οι ιδιότητες πήξης των εναλλακτικών πρωτεϊνών επηρεάζονται από διάφορους παράγοντες, όπως ο τύπος της πρωτεΐνης, η περιεκτικότητα σε πρωτεΐνη, η θερμοκρασία, η τιμή του pH, η ισχύς Ionic και άλλα συστατικά.
Συμπέρασμα
Οι ιδιότητες πηκτωματοποίησης που προκαλούνται από τη θερμότητα δύο εναλλακτικών πηγών πρωτεϊνών (πρωτεΐνες φυτικής προέλευσης και μη φυτικής προέλευσης νέες πρωτεΐνες) μελετήθηκαν με την εφαρμογή περιστροφικής ρεομετρίας. Οι καμπύλες ζελατινοποίησης του μέτρου αποθήκευσης, G', και του μέτρου απώλειας, G'', καταγράφηκαν και ερμηνεύτηκαν κατά τη διάρκεια της σάρωσης θερμοκρασίας. Η θερμοκρασία πήξης, η σταθερότητα πηκτής και η αντοχή πηκτής των εναλλακτικών πρωτεϊνών αναλύθηκαν και συγκρίθηκαν με εκείνες των ζωικών πρωτεϊνών (αυγό). Οι μετρήσεις αυτές είναι γρήγορες και απαιτούν σχετικά small ποσότητα αντιπροσωπευτικού δείγματος.