Introduction
Avec l'augmentation de la population mondiale et la demande croissante de protéines, certaines sources de protéines alternatives ont récemment suscité un intérêt croissant, telles que les protéines végétales, les protéines de viande cultivées, les protéines issues de la fermentation, les protéines d'insectes comestibles et les algues [1]. La propriété gélifiante est l'une des fonctions les plus importantes des protéines alternatives, car elle contribue à la texture et au goût des produits alimentaires. La gélification se produit au cours de la transformation et de la fabrication des produits alimentaires. Le chauffage est l'une des méthodes les plus fréquemment utilisées pour former des gels avec les protéines alternatives. Après dénaturation et dépliage des molécules de protéines par chauffage et enveloppement d'eau, celles-ci s'agrègent pour former une structure de réseau tridimensionnelle, c'est-à-dire la structure du gel.
Dans la pratique, un rhéomètre rotatif permet d'étudier les propriétés de gélification des protéines alternatives induites par la chaleur, telles que la température de gélification, la stabilité et la résistance du gel.
Matériaux et conditions de mesure
Les poudres de protéines ont été dispersées dans de l'eau déminéralisée à des concentrations de protéines définies (concentré de protéines de pois : 10 % en poids et 7 % en poids, protéines d'insectes : 10 % en poids). Les suspensions de protéines ont été agitées à l'aide d'un agitateur magnétique à température ambiante pendant 2 heures. Outre l'échantillon d'œuf entier, un échantillon de blanc d'œuf a été préparé en enlevant le jaune d'œuf et en fouettant vigoureusement à température ambiante pendant quelques minutes pour obtenir une solution homogène.
Un rhéomètre NETZSCH Kinexus Prime pro+ , équipé d'un système plaque-plaque (diamètre : 40 mm, espace : 0,5 mm) a été utilisé pour effectuer les mesures sur les échantillons de concentré de protéines de pois, d'œuf entier et de blanc d'œuf. Un balayage de température a été effectué en augmentant la température de 25°C à 95°C à une vitesse de 5°C/min. Après avoir atteint la température la plus élevée, les gels induits par la chaleur ont été maintenus pendant 10 minutes pour étudier la stabilité du gel. Le module de stockage (G') et le Module visqueuxLe module complexe (composante visqueuse), module de perte ou G'', est la partie "imaginaire" du module complexe global des échantillons. Cette composante visqueuse indique la réponse liquide ou déphasée de l'échantillon mesuré. module de perte (G'') ont été enregistrés pendant l'expérience. Les résultats obtenus pour les protéines végétales et les protéines d'œuf ont été comparés à ceux obtenus pour les protéines d'insectes [2].

Au milieu : Œufs frais (utilisés comme protéines animales pour la comparaison) : l'œuf entier et le blanc d'œuf ont été analysés, respectivement.
À droite : Nouvelles protéines non végétales : protéines d'insectes (teneur en protéines brutes de 68,7 g/100 g) extraites de pupes d'insectes de la mouche soldat noire (BSF) dans une étude précédente [2]
Résultats des mesures et discussion
Les propriétés gélifiantes de différentes protéines alternatives et d'une source de protéines animales typique (œuf) ont été étudiées pendant et après le chauffage à l'aide de la rhéométrie rotative.
La figure 2 montre le module de stockage, G', et le Module visqueuxLe module complexe (composante visqueuse), module de perte ou G'', est la partie "imaginaire" du module complexe global des échantillons. Cette composante visqueuse indique la réponse liquide ou déphasée de l'échantillon mesuré. module de perte, G'', du gel formé avec le concentré de protéines de pois pendant le traitement thermique. Une augmentation de G' et G'' se produit lorsque la température augmente jusqu'à 55°C environ. Cela est dû à la dénaturation des protéines. Après le traitement thermique, en augmentant encore la température, G' est plus élevé que G'' à une concentration de protéines de 10 % en poids, ce qui montre un comportement de gel solide.
En outre, une expérience sur le concentré de protéines de pois à une concentration de protéines plus faible de 7 % en poids indique que G'' est plus élevé que G' avec l'augmentation de la température de chauffage, ce qui suggère un comportement de gel liquide faible.
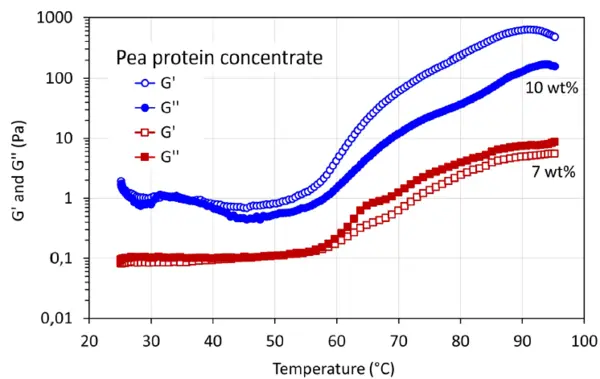
Cependant, le croisement G' = G'' n'a pas été observé dans cette étude sur le concentré de protéines de pois.
Dans une étude précédente sur la gélification induite par la chaleur avec les protéines d'insectes de la chrysalide BSF [2], il a été constaté qu'en augmentant la température au-dessus de 50°C, G' et G'' augmentaient tous les deux, en raison de la dénaturation de la protéine. L'échantillon étudié a commencé à former un gel, ce qui a été indiqué par le croisement G' = G'' à 60°C, la température du point de gélification.
La progression de la courbe en fonction de la température pour les protéines d'insectes est différente de celle du concentré de protéines de pois. Ces différences de comportement en matière de gélification peuvent être attribuées aux différentes compositions des matériaux et aux caractéristiques individuelles des protéines, telles que les différents acides aminés hydrophiles et hydrophobes possibles et leurs ratios parmi les différentes protéines alternatives.
Les courbes de gélification des échantillons d'œuf entier et de blanc d'œuf présentent la transition sol-gel typique pendant le balayage de température. À partir d'environ 60°C, une augmentation significative de G' et G" peut être observée, ce qui peut s'expliquer, par exemple, par des changements structurels ou la dénaturation des protéines. La figure 3 illustre G' et G'' du gel formé avec la solution d'œuf entier pendant le traitement thermique. G' présente une augmentation apparente à environ 62°C et une forte augmentation à environ 75°C, tandis que G'' présente une augmentation spectaculaire à environ 75°C. Le point de croisement se produit à environ 74°C. Pour l'échantillon de blanc d'œuf (figure 4), G' et G'' présentent deux augmentations apparentes à environ 64°C et 75°C, respectivement. Le point de croisement se situe à environ 62,5°C. Les phénomènes de dénaturation observés peuvent être liés à la composition chimique de l'échantillon d'œuf entier (blanc et jaune d'œuf) et de l'échantillon de blanc d'œuf.
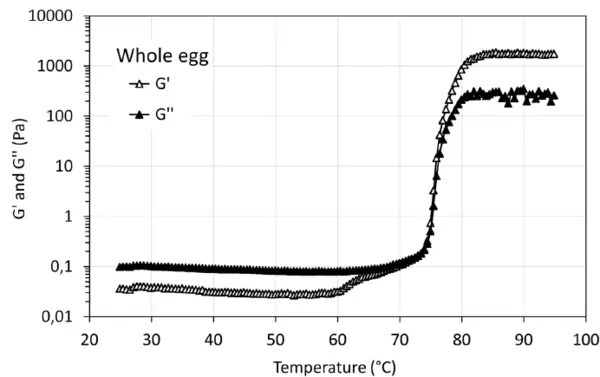
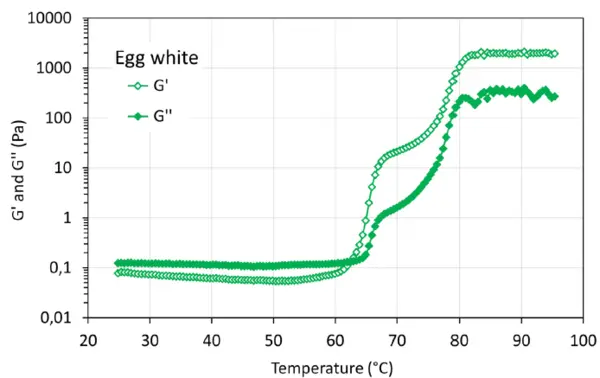
La figure 5 illustre la force du gel, G', et la stabilité dans un temps de maintien de 10 minutes après avoir atteint la température la plus élevée. Les gels induits par la chaleur à partir de protéines d'œuf présentent la force la plus élevée et sont très stables. Cette propriété de gel stable a également été observée à une température supérieure à 85°C dans les figures 3 et 4. Pour l'échantillon de concentré de protéines de pois à 10 % en poids, il faut environ 4 minutes pour atteindre la force de gel la plus élevée, puis le gel formé est stable, alors que la force de gel diminue légèrement pour l'échantillon de concentré de protéines de pois à 7 % en poids. Cela peut être dû à la déformation (destruction) de la structure de gel faible formée pendant la mesure. Par rapport à la force du gel formé avec des protéines d'insectes [2], ces gels présentent des forces de gel différentes, G', dans l'ordre suivant
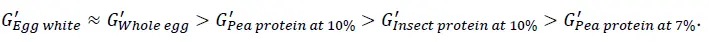
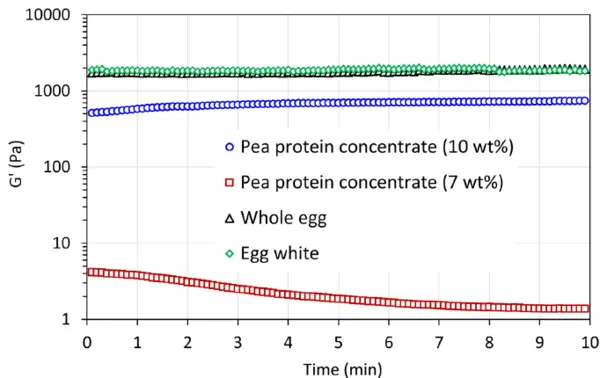
Cela pourrait suggérer que les différentes protéines alternatives ont des applications potentielles différentes. Par exemple, les gels induits par la chaleur avec une valeur G' plus faible ou un réseau de gel faible pourraient être intéressants et adaptés aux formulations alimentaires liquides telles que les boissons à base de plantes ou les laits alternatifs, tandis que les gels avec une valeur G' plus élevée ou un réseau de gel fort seraient intéressants pour les analogues de produits laitiers et de viande, etc.
Il convient de mentionner que les propriétés gélifiantes des protéines alternatives sont influencées par différents facteurs, tels que le type de protéine, la teneur en protéines, la température, la valeur pH, la force Ionic et d'autres composants.
Conclusion
Les propriétés de gélification induites par la chaleur de deux sources de protéines alternatives (protéines d'origine végétale et nouvelles protéines d'origine non végétale) ont été étudiées en appliquant la rhéométrie rotationnelle. Les courbes de gélification du module de stockage, G', et du Module visqueuxLe module complexe (composante visqueuse), module de perte ou G'', est la partie "imaginaire" du module complexe global des échantillons. Cette composante visqueuse indique la réponse liquide ou déphasée de l'échantillon mesuré. module de perte, G'', ont été enregistrées et interprétées pendant le balayage de température. La température de gélification, la stabilité et la force de gélification des protéines alternatives ont été analysées et comparées à celles des protéines animales (œuf). Ces mesures sont rapides et nécessitent une quantité relativement importante d'échantillon représentatif ( small ).